
减数分裂是真核生物有性生殖所必需的一种特殊的细胞分裂方式,最终产生单倍体配子。减数分裂重组是减数分裂的核心事件,其起始于保守的DNA拓扑异构酶复合体(SPO11-1/SPO11-2/MTOPVIB)介导DNA双链断裂(DSB)的产生。减数分裂同源染色体重组除了创造新的等位基因组合,既确保了生物世代间遗传物质在稳定传递,又增加了后代的遗传多样性。在酵母、哺乳动物和植物的研究中发现减数分裂DSB沿染色体的分布并非随机,会优先发生在一些小区域(~1-2kb),被称为DSB热区,其具有显著的表观遗传修饰和基因组特征。植物中发现组蛋白修饰,如H3K4me3、H3K36me3、组蛋白变体H2A.Z等在热区富集。出芽酵母和植物中都表明DSB倾向发生在基因的转录起始位点(TSS),伴随着低核小体密度,暗示了DSB通常发生在具有转录活跃特征的染色体开放区域。有趣的是,DSB位点上的基因转录和DNA修复呈现负相关,而植物减数分裂过程中DSB位点上的基因转录状态及其调控机制还未有报道。
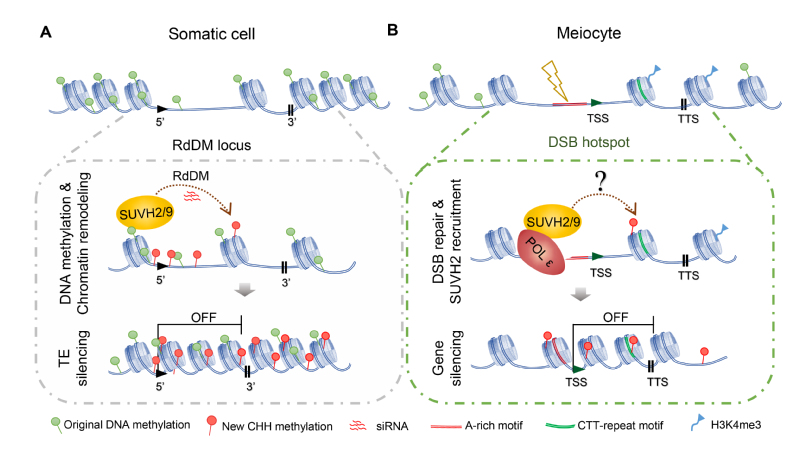
图1. POL ε-SUVH2/9调控减数分裂DSB偶联的基因沉默的模型
2022年10月3日,王应祥课题组在PNAS在线发表了题为“DNA polymerase epsilon interacts with SUVH2/9 to repress the expression of genes associated with meiotic DSB hotspot in Arabidopsis ” 的研究论文。该研究揭示了植物减数分裂过程中DNA双链断裂(DSB)位点基因沉默的分子机制,揭示了DNA聚合酶epsilon(POL ε)和SUVH2/9在减数分裂细胞中不同于体细胞中的新功能。
研究者在前期工作中发现了不同DNA聚合酶(Pol α、Pol δ和Pol ε)在减数分裂重组DNA修复环节中作用,其中Pol ε的催化亚基POL2A对减数分裂DSB的修复和重组分布调控至关重要(Huang et al., PNAS, 2015)。然后,细胞进入在进一步探索中,DNA聚合酶仍旧会分布在整个染色体上,本研究报道了其在常染色质DSB富集区域的功能。研究者发现POL2A和RdDM(RNA-directed DNA methylation)的组分SUVH2相互作用。进一步研究验证了POL2A的N端和SUVH2/9的SET结构域互作,运用N-SIM (Structured Illumination Microscopy)观察到SUVH2在减数分裂时期与γ-H2AX标记的DSB共定位,并且该定位依赖于POL2A。暗示着POL2A在减数分裂DSB修复过程中招募SUVH2。
RdDM途径中SUVH2和其同源蛋白SUVH9影响靶位点小RNA的产生和CHH(H表示A、T或G)甲基化。通过高通量测序,研究者发现有别于SUVH2/9在体细胞中抑制TE表达的作用,SUVH2/9和POL2A在减数分裂细胞中共同抑制865个基因的表达。并且这865个基因不直接参与调控减数分裂功能,但在其转录起始位点(TSS)区域具有明显的DSB热区的特征,包括低核小体密度,A-rich基序和SPO11-1的富集等。这一结果暗示了POL2A和SUVH2/9可能在减数分裂DSB位置发挥着基因沉默的作用。同时研究者发现这一功能并不类似于体细胞中通过调控DNA的甲基化或小RNA来实现,但POL2A和SUVH2/9都影响了减数分裂染色体的浓缩,表明它们可能直接通过调控染色质结构从而抑制基因表达(图1)。
最后研究者通过遗传分析还发现了SUVH2/9的突变会进一步加强POL2A突变体的减数分裂DSB修复异常和雄性败育的表型。并且在各突变体中通常上调表达基因越多伴随着减数分裂DSB修复异常的缺陷越严重。因此我们推测POL2A和SUVH2/9在DSB位点诱导的基因沉默对正常减数分裂DSB修复具有重要意义。
综上所述,该研究揭示了植物DSB位置的基因转录状态,发现了DNA聚合酶epsilon和SUVH2/9共同调控减数分裂DSB偶联的转录沉默的新机制(图1),该机制可能对于减数分裂过程的DSB修复和基因组稳定性至关重要。
王应祥教授课题组博士后王聪为该论文的第一作者,王聪博士在研究生和博士后期间以第一作者(含共同)在PNAS(2篇)、Plant Cell(1篇)和Plant Physiology(2篇)发表5篇论文。王应祥教授和尤辰江青年研究员为共同通讯作者。
文章链接:https://www.pnas.org/doi/full/10.1073/pnas.2208441119
